하플로그룹 O1b2 (Y-DNA)
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
하플로그룹 O1b2 (Y-DNA)는 동북아시아에서 주로 발견되는 Y-DNA 유전 형질로, 현대 일본인과 한민족 남성에게서 높은 빈도로 나타난다. 일본 열도 또는 한반도에서 기원한 것으로 추정되며, O1b2의 하위 그룹인 O-K10은 현대 일본인과 한국인에게서 많이 발견된다. O1b2는 야요이 시대의 인골에서 발견되며, 벼농사 문화를 전파한 야요이인과 관련이 있을 것으로 추정된다.

더 읽어볼만한 페이지
- 야요이 시대 - 요시노가리 유적
요시노가리 유적은 일본 사가현에 있는 야요이 시대의 대규모 마을 유적으로, 정교한 방어 시설과 다양한 유구, 풍부한 유물을 통해 당시 사회상과 주변 지역과의 활발한 교류를 보여주는 중요한 유적이며 현재는 역사공원으로 조성되어 있다. - 야요이 시대 - 곡옥
곡옥은 동물의 이빨이나 태아 모양을 본뜬 장신구로, 일본에서 유래하여 권력의 상징으로 사용되었으며, 현재는 부적이나 장식품으로 쓰인다. - 인류 Y-DNA 하플로그룹 - 하플로그룹 K (Y-DNA)
하플로그룹 K는 약 47,000~50,000년 전 중동 또는 중앙아시아에서 기원한 것으로 추정되는 Y-염색체 DNA 하플로그룹으로, 하위 계통인 L, T, NO, M, S, P 등을 통해 유라시아, 오세아니아, 아메리카 대륙 등 전 세계에 분포한다. - 인류 Y-DNA 하플로그룹 - Y염색체 아담
Y염색체 아담은 현존하는 모든 인류의 부계 혈통을 따라 올라갈 때 만나는 가장 최근의 공통 조상을 지칭하는 비유적인 용어이며, 단 한 명의 남성이 아닌 Y염색체에 돌연변이가 발생하기 이전의 남성 집단을 의미한다.
2. 분포
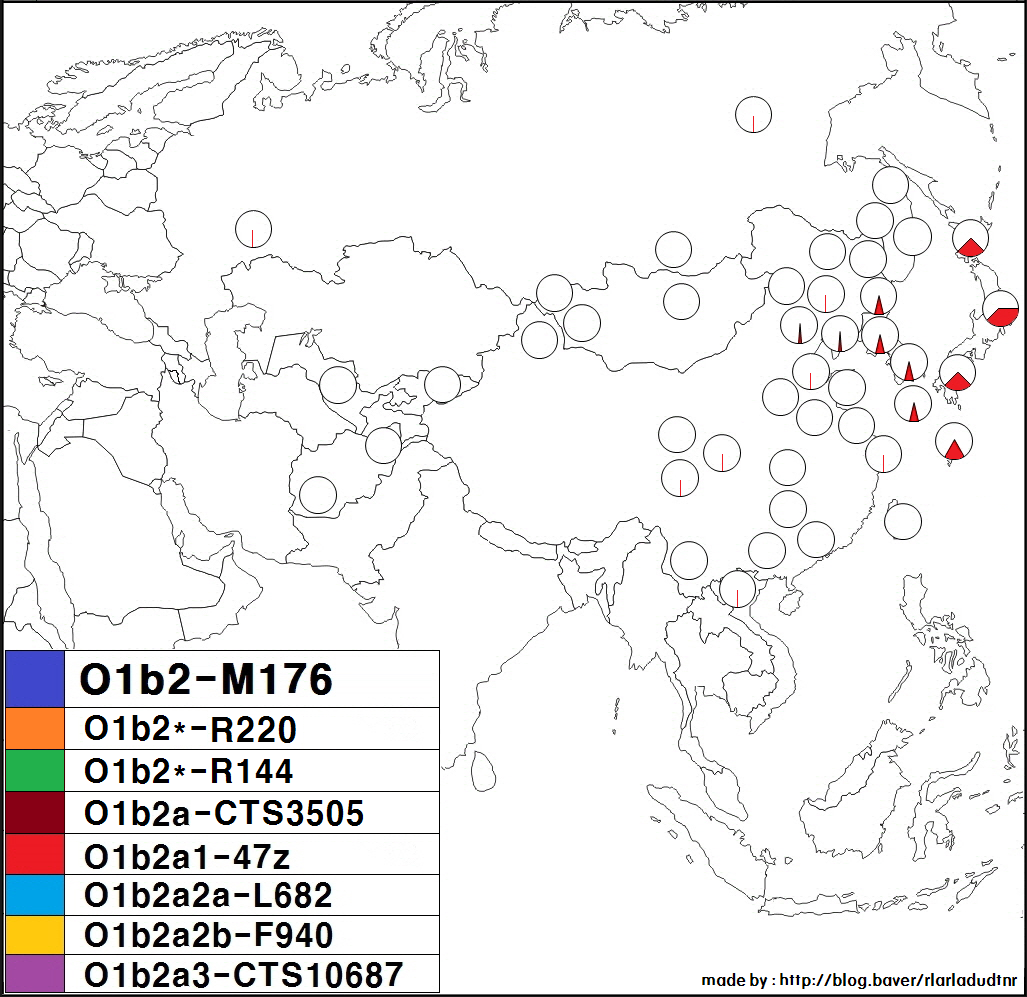
하플로그룹 O1b2는 동북아시아에서 주로 나타나는 Y-DNA 유전 형질로, 서쪽으로는 몽골 서부의 알타이산맥과 자흐친부터 동쪽으로는 일본 및 오키나와에 이르는 지역에서 주로 발견된다.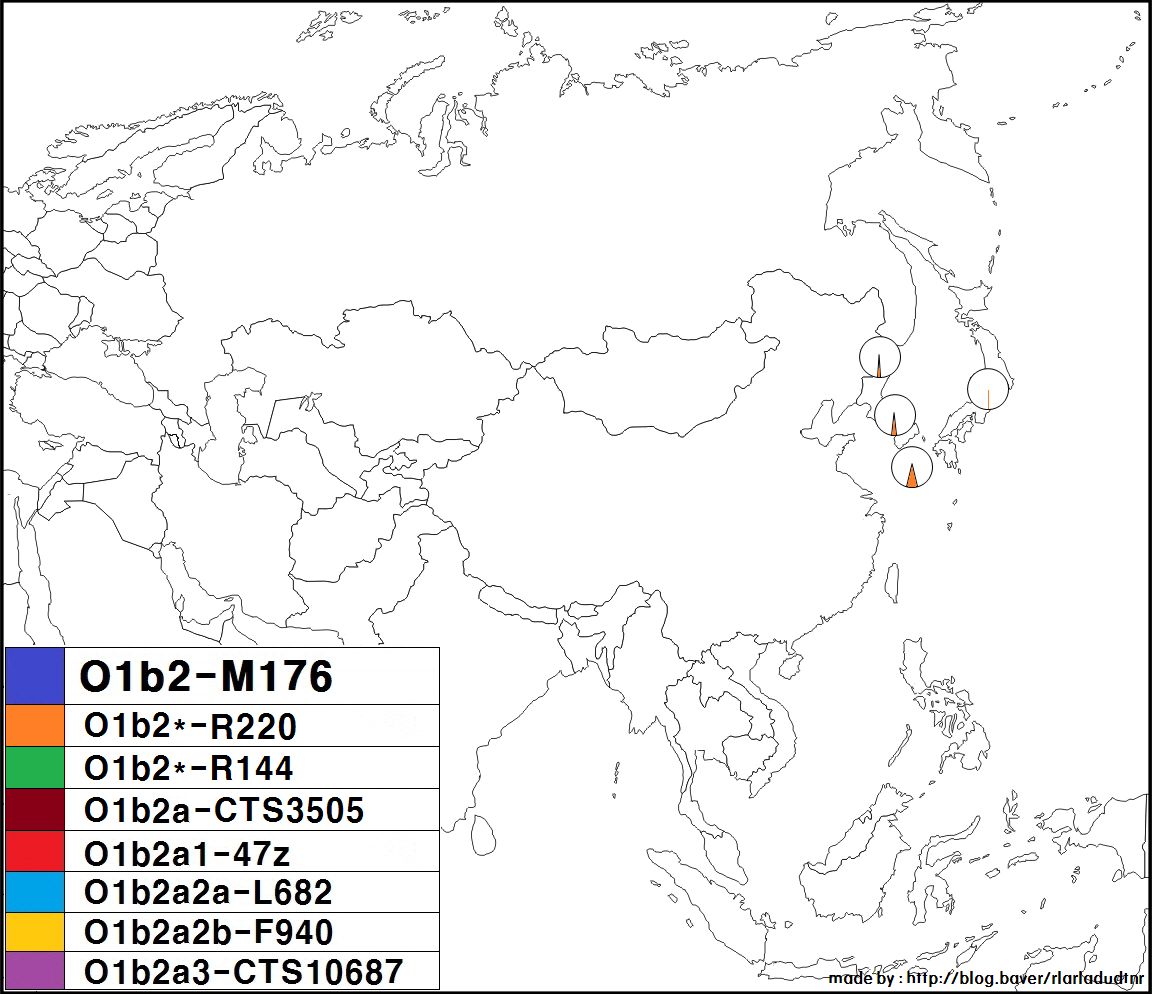
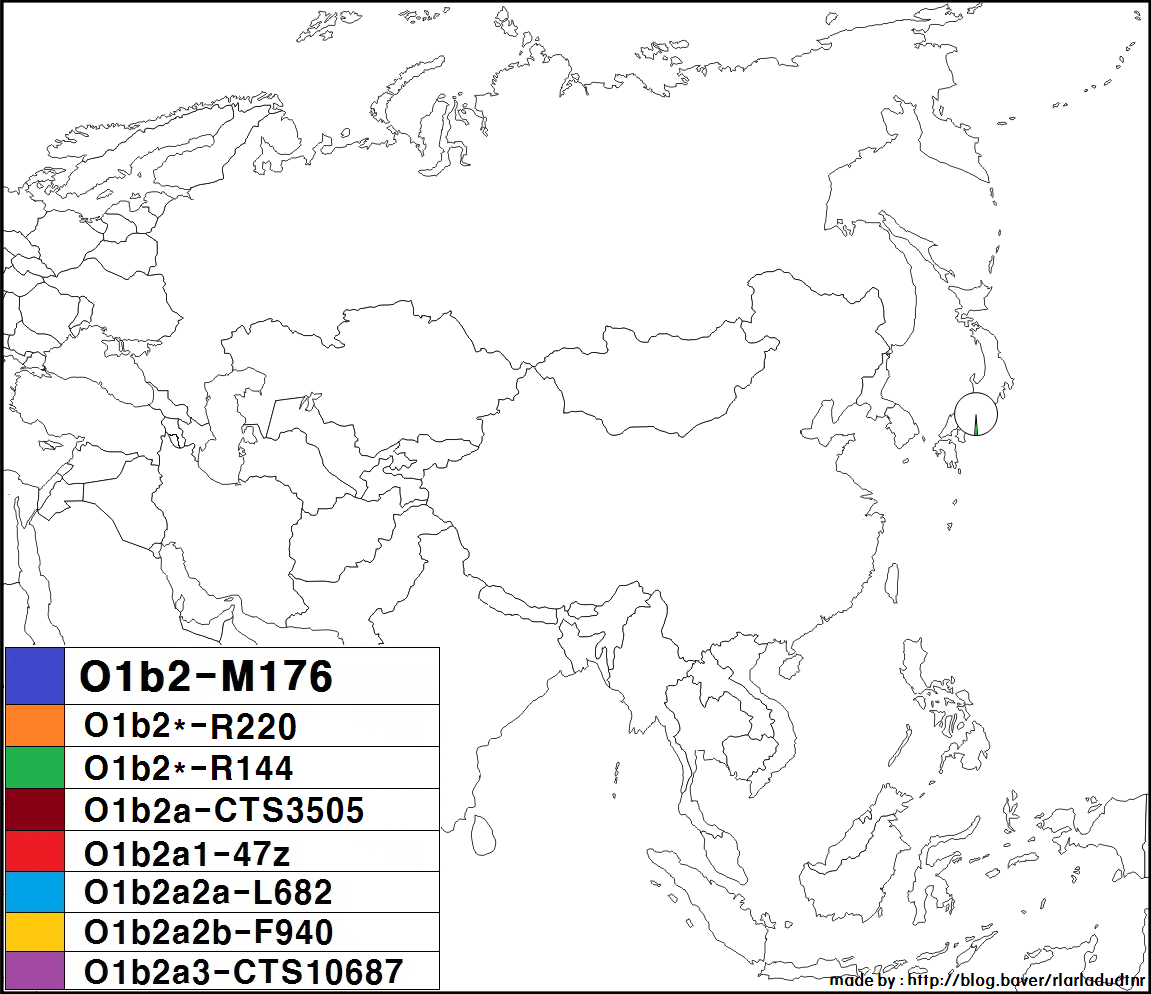
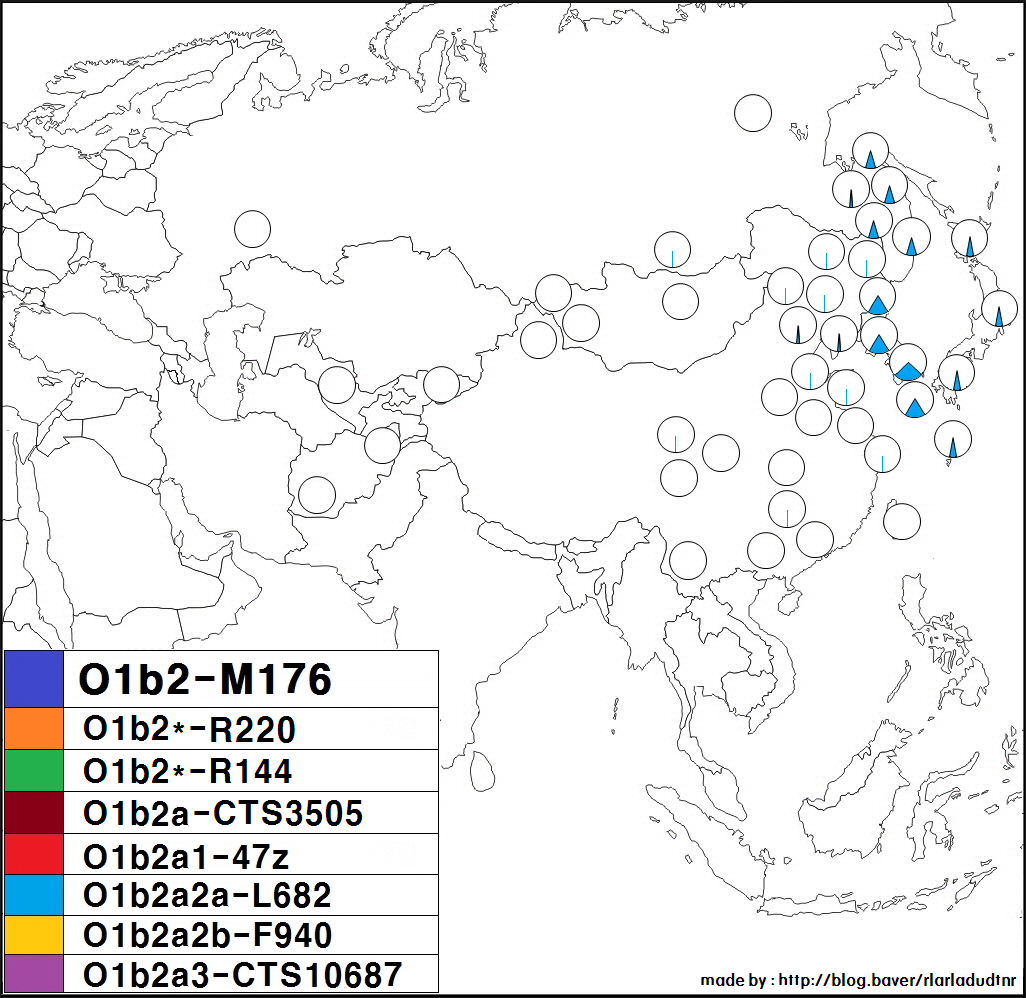
일본 국내, 한국 국내의 각 지역 분포에서는 특히 편향성은 보이지 않는다. 다만, 하위 그룹에 관해서 말하자면, 일본인은 O-CTS713이 많은 반면, 한민족은 O-CTS723이 많다. O-CTS10687은 일본이나 한국 등에서 드물게 발견된다.
아이누 민족에게서는 발견되지 않으며 (16명의 샘플 중 0명[13], 4명의 샘플 중 0명[1]), 야요이 시대 이후의 왜인(야요이인)의 유전자일 것으로 추정된다.[14] 동아시아 북동부에 수도 농경을 가져온 집단이라는 견해가 있다.
만주족에서는 23mofang의 통계에 따르면 약 3.10% (91/2938)의 현대 만주족 남성에게 관찰되고 있다.
현대 만주족 남성의 약 1할이 한국계라고 한다면 쉽게 설명이 되는 양상을 띠고 있다.[1]
몽골에 관해서는, 중화인민공화국 내에 거주하는 소위 몽골족은 23mofang의 통계에 따르면 약 1.51% (23/1521)의 몽골족 남성이 하플로그룹 O-F1204/O-K10에 속해 있다.[15][16]
몽골 국내를 대상으로 한 많은 연구 논문에서는 하플로그룹 O1b2에 속할 가능성이 있는 남성이 전무하다.
몽골국 및 중국 "몽골족"의 샘플 중에서 하플로그룹 O1b2에 속하는 사람이 관찰된 예는 다음과 같다.
- Di Cristofaro 등 (2013년)의 논문[24] 몽골국 거주 몽골인 총 1/160 = 0.6% O2b-M176
- * Mongol-NorthEast 1/20 O2b-M176
- * Mongol-SouthWest 0/2 O2b-M176
- * Mongol-Central 0/18 O2b-M176
- * Mongol-SouthEast 0/23 O2b-M176
- * Mongol-NorthWest 0/97 O2b-M176
- Xuwei Hou 등 (2022년)의 논문[25]
- * 랴오닝성 푸신시 몽골족 1/13 O1b2a1a1-K7/CTS11723
- 허광린 등 (2023년)의 논문[26] (중국 몽골족 5/175 = 2.86% O1b2a1a)
- * 산둥성 몽골족 1/6 O1b2a1a1
- * 랴오닝성 푸신시 몽골족 1/9 O1b2a1a1
- * 헤이룽장성 몽골족 1/10 O1b2a1a3a
- * 랴오닝성 차오양시 몽골족 1/12 O1b2a1a1
- * 내몽골 자치구 츠펑시 몽골족 1/22 O1b2a1a1
하플로그룹 O1b2에 속하는 몽골 민족의 사람이 주로 몽골 민족 거주 지역의 동부, 특히 남동부의 사람이라는 것을 알 수 있다. 예외로, 가토 토오루 등에 의한 2005년의 논문에서는, 하플로그룹 O1b2가, 할하 (몽골국의 주류 부족) 및 호톤 (이슬람교도)의 샘플에서는 검출되지 않은 반면, 우리양하이 및 자흐친 (둘 다 몽골국 서부에 사는 부족)의 샘플에서는 어느 정도 검출되었다고 한다.[27]
ISOGG영어(ver.12.244) 2017년 9월 10일 조사에 따른 하플로그룹 O1b2의 하위 트리는 다음과 같다.[1]
또한, 한국의 한 연구팀에 의한 일련의 논문에서는 하리하라 신지로부터 제공받았다고 하는[28] "동몽골의 부랴트족"의 샘플을 다루고 있으며 49명 중 1명[1] 또는 50명 중 1명[1]에게 하플로그룹 O1b2-M176(x47z)이 관찰되었다고 한다.
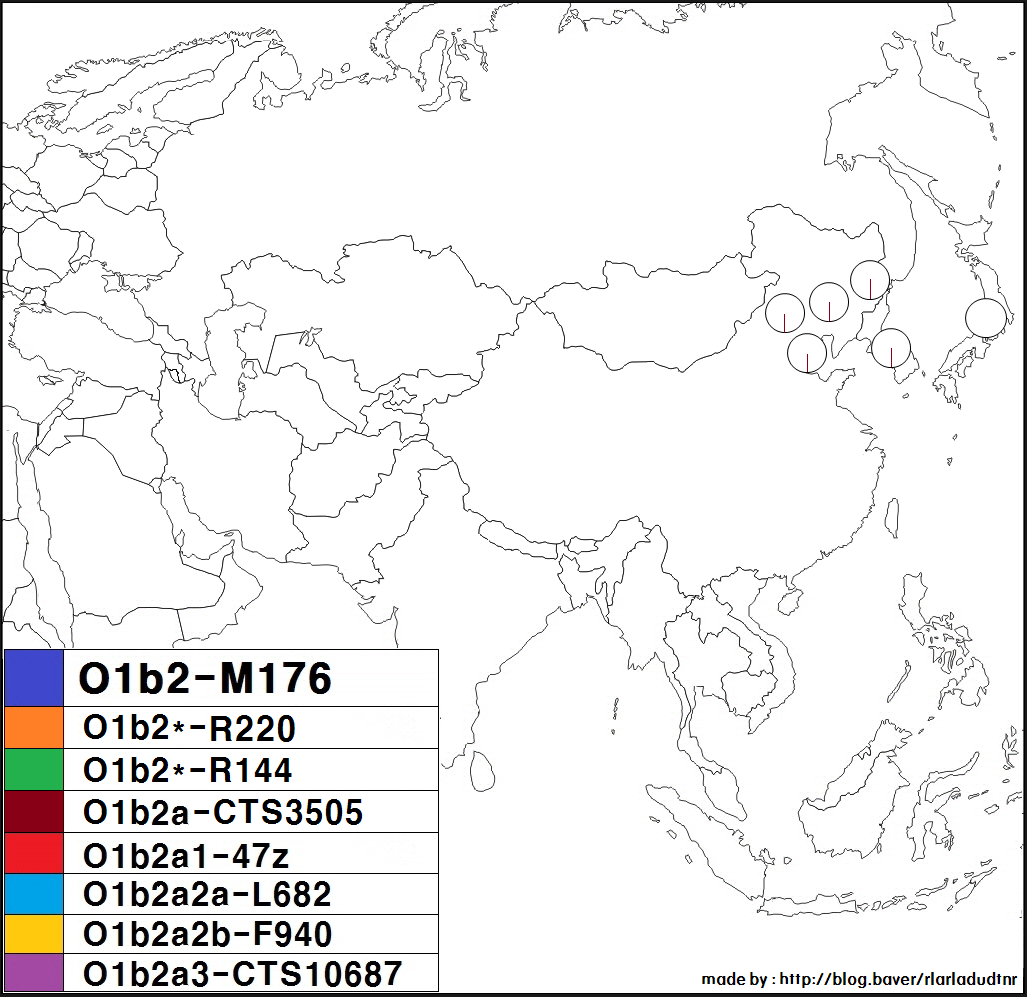
또한, 러시아 국내에 거주하는 우데게[29], 그리고 중국 국내에 거주하는 다우르족, 나나이족, 에벤키족, 시버족, 한족, 회족, 위구르족, 투자족, 쓰촨성 간쯔 티베트족 자치주 신룽현의 티베트족 (캄파) 등에서도 드물게 관찰된 예가 있다.
O-K10(F1204)에 속하지 않는 O-M176은 일본인, 한국인, 만주족, 한족 등에서 낮은 빈도로 검출되고 있다.
3. 계통
'''O1b2a1a''' (F3356)
* '''O1b2a1a1''' (CTS713, CTS11986): 일본 열도(Hammer 2006) 혹은 한반도에서 유래하였다.
** '''O1b2a1a1a''' (CTS1875): 일본 열도에서 고빈도로 나타난다.
*** '''O1b2a1a1a1''' (CTS10682): 일본 열도
** '''O1b2a1a1b''' (Z24598): 일본 열도
** '''O1b2a1a1c''' (CTS203): 일본 열도
* '''O1b2a1a2''' (F2868, F3110, K4): 일본 열도
** '''O1b2a1a2a''' (L682): 한반도 혹은 만주 지역에서 유래하였다. 한국인 20%, 오키나와인 12%, 우데게족 10%, 나나이족 8%, 일본인 6%, 만주족 3%에게서 나타난다.
*** '''O1b2a1a2a1''' (CTS723): 한반도
'''O1b2a1a2a1a''' (CTS7620): 일본 열도, 나나이족
'''O1b2a1a2a1b''' (A12446)
* '''O1b2a1a2a1b1''' (PH40)
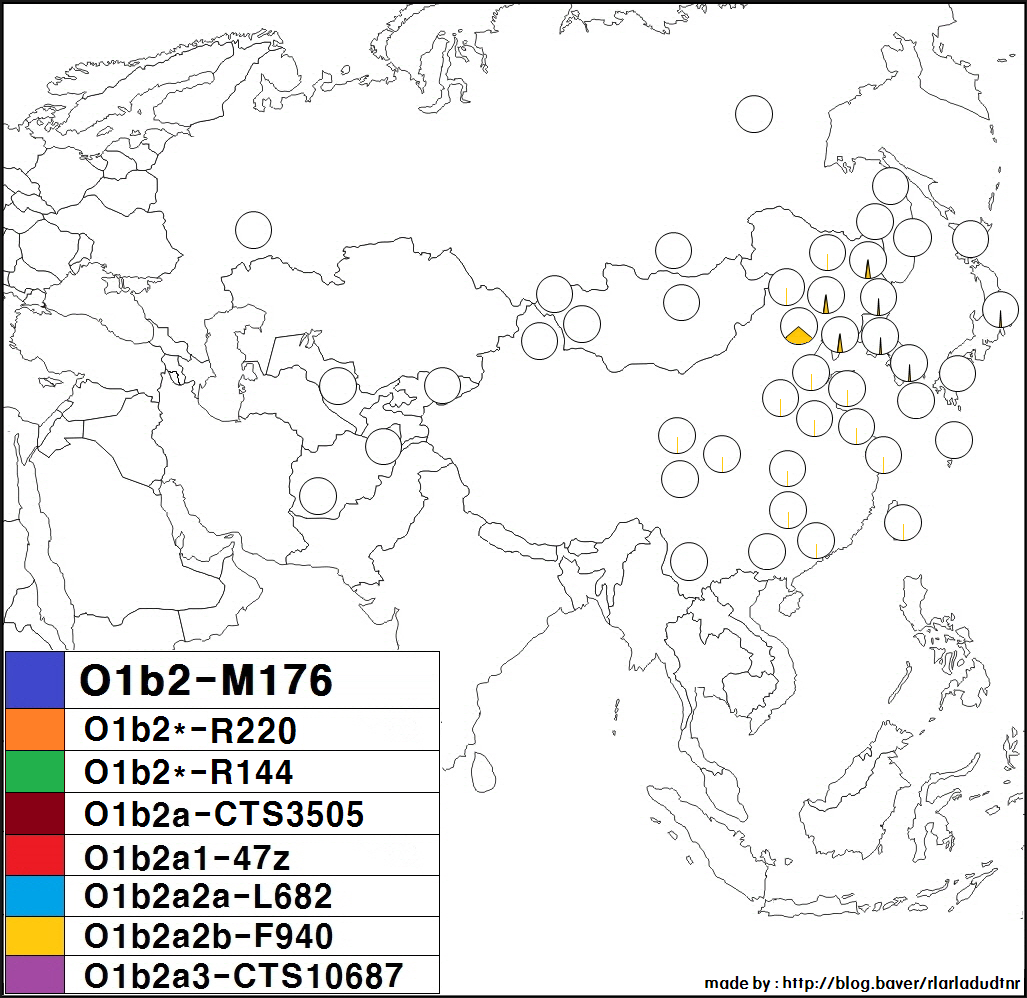
** '''O1b2a1a2b''' (F940, F1912, F3390): 한반도와 만주 지역에서 유래하였다. 만주족에게서 0-26%로 가장 고빈도로 나타난다.
* '''O1b2a1a3''' (CTS10687, F1800): 강원도에서 4%로 가장 고빈도로 발견된다. 그 외 주로 북방 소수민족에서 소수로 발견된다. (시버족, 부랴트족, 우랑카이인(북서몽골 지역), 자흐친족(남서몽골 지역), 키르키즈스탄, 우즈베키스탄, 아프가니스탄(바다흐샨 지역)).
** '''O1b2a1a3a''' (CTS1215)
'''O1b2a1b''' (CTS562, CTS562*): 중국(베이징)
가장 가까운 하플로그룹 O1b1과의 분기는 약 31,108년 전 (95% CI 22,844~34,893)[3]으로 여겨진다. 가장 가까운 공통 조상은 25,800년 전 (95% CI 28,400~23,300)[1] 또는 26,650년 전 (99% CI 33,045 - 21,160)[1], 혹은 25,450년 전[4]까지 거슬러 올라간다고 추정된다.
4. 역사
하플로그룹 O1b1에서 약 31,108년 전 (95% CI 22,844~34,893) 분기된 것으로 추정된다.[3] 가장 가까운 공통 조상은 25,800년 전 (95% CI 28,400~23,300)[1], 26,650년 전 (99% CI 33,045 - 21,160)[1], 혹은 25,450년 전[4]으로 거슬러 올라간다.
O-M175의 하위 그룹 중 하나로, 벼농사 문화를 전파한 야요이인(왜인)으로 추정된다.[5] 현대 일본인의 약 32%가 O-M176의 하위 그룹에 속하는 Y-DNA를 가지고 있다.[34][1]
2024년 9월 현재까지 하플로그룹 O1b2로 판정된 Y-DNA는 다음과 같다.
사키야 미쓰루는 벼 품종을 근거로 O-M176 그룹이 2800년 전 중국강남에서 산둥반도, 일본 열도, 한반도로 수도 재배를 가져왔다고 주장했지만, 중국강남에서 O-M176이 거의 발견되지 않아 신빙성이 낮다.[1] O-M176은 현대 일본 및 한국·조선에 높은 빈도로 나타나며, 다른 지역 기원 증거는 아직 없다.
조몬 시대 말기부터 야요이 시대에 걸쳐 널리 보이는 각목돌대문 토기와 비슷한 토기가 연해주 남서부 시니 가이 문화에서 발견[156]되면서, 각목돌대문 토기 및 하플로그룹 O1b2에 속하는 야요이인 기원을 연해주 남서부에서 찾는 시각이 있다.
아오야카미데라지 유적 야요이 시대 인골 Y-DNA 분석 결과, 10개체 중 2개체가 O1b2a1에 속했고, 그 중 1개체는 현대 일본인에게서 많이 보이는 O1b2a1a1에 속했다. 다른 개체는 하플로그룹 D 3개체, C1a1 2개체, O (하위 계통 미발표) 1개체였고, 2개체는 특정 불가능했다.[160][157]
나가사키현 사세보시 시모모토야마 이와카게 유적 곽식 석관 출토 야요이 시대 남성 인골 (시모모토야마 3호, 방사성 탄소 측정 연대 2001–1931 cal BP [1σ])도 O1b2a1a1-CTS713에 속한다는 결과가 발표되었다.[158][159]
위 연구 결과에 따라, O1b2는 야요이 시대 이나바 국 및 히젠 국 지역에, C1a1과 D는 이나바 국 지역에 이미 존재했음이 보고되었다.[160] 현대 일본인 남성 70% 이상이 D, O1b2, C1a1 중 하나에 속하므로, 야요이 시대 서일본 주민과 현대 일본인 사이에는 Y 염색체 (남계) 기준 큰 차이가 없다. (단, 현대 일본인 남성 약 20%를 차지하는 O2는 2023년 현재 야요이 시대 유적에서 발견되지 않았다.)
5. 유적
2024년 9월 현재, 하플로그룹 O1b2에 속한다고 판정된 Y-DNA는 다음과 같은 유적에서 검출되었다.[6][1][7][8][9]
아오야카미지 유적에서 출토된 야요이 시대 인골 중 Y-DNA가 분석된 10개체 중 2개체가 하플로그룹 O1b2a1에 속했고, 그 중 1개체는 현대 일본인에게서 많이 보이는 O1b2a1a1에 속했다.[160][157] 나가사키현 사세보시에 있는 시모모토야마 이와카게 유적의 곽식 석관에서 출토된 시모모토야마 3호라는 야요이 시대 남성 인골(방사성 탄소 측정에 의해 연대는 2001–1931 cal BP [1σ]로 추정)도 하플로그룹 O (TheYtree에 따르면 O1b2a1a1-CTS713[158])에 속한다는 결과가 발표되었다.[159]
이러한 연구 결과에 따라, 하플로그룹 O1b2가 야요이 시대에는 이미 이나바 국 및 히젠 국 지역에 존재했음이 보고되었다.[160]
6. 현대 한국 사회에 대한 시사점
하플로그룹 O1b2는 현대 일본인과 한민족 남성에게서 약 30% 정도로 발견된다.[10] 일본과 한국 내 지역별 분포에서는 큰 편향성은 보이지 않지만, 하위 그룹에서는 차이가 나타난다. 일본인은 O-CTS713이 많은 반면, 한국인은 O-CTS723이 더 많이 발견된다.[1]
만주족의 경우, 23mofang 통계에 따르면 약 3.10% (91/2938)의 남성에게서 O1b2가 관찰된다.[1] 하위 그룹별 비율은 현대 한민족과 거의 유사하며, 이는 현대 만주족 남성 중 약 10%가 한국계일 가능성을 시사한다.[1]
몽골족의 경우, 중화인민공화국 내 몽골족은 23mofang 통계에서 약 1.51%(23/1521)의 남성이 O-F1204/O-K10에 속한다.[15][16] 그러나 몽골 국내 연구에서는 O1b2에 속하는 남성이 거의 발견되지 않거나 매우 드물게 발견된다.[17][1][18][19][20][21][22][23] O1b2가 발견되는 몽골인은 주로 몽골 민족 거주 지역의 동부, 특히 남동부에 집중되는 경향을 보인다.[24][25][26]
이 외에도 러시아의 우데게족[52][53][54], 중국의 다우르족, 나나이족[47][48][49][50], 에벤키족[1], 시버족[1] 등에서도 드물게 O1b2가 관찰된다.
참조
[1]
웹사이트
https://isogg.org/tr[...]
[2]
웹사이트
ISOGG系統樹
http://www.isogg.org[...]
[3]
논문
Episodes of Diversification and Isolation in Island Southeast Asian and Near Oceanian Male Lineages.
[4]
웹사이트
TheYTreeによるO1b2-P49の系統樹及び最も近い共通祖先の推定年代
https://www.theytree[...]
[5]
논문
Munda languages are father tongues, but Japanese and Korean are not
https://www.cambridg[...]
[6]
간행물
鳥取県鳥取市青谷上寺地遺跡出土弥生後期人骨の核DNA分析
国立歴史民俗博物館
2021-03
[7]
논문
Northeastern Asian and Jomon-related genetic structure in the Three Kingdoms period of Gimhae, Korea
https://doi.org/10.1[...]
2022
[8]
논문
Genomic detection of a secondary family burial in a single jar coffin in early Medieval Korea
https://doi.org/10.1[...]
[9]
논문
"[調査研究活動報告] 島根県出雲市猪目洞窟遺跡出土人骨の核DNA分析"
https://rekihaku.rep[...]
国立歴史民俗博物館
[10]
문서
ただし、朝鮮半島及び満州地域に多いO1b2a1a2a-L682と日本人に多いO1b2a1a1-47Z/K7の最も近い共通祖先は、約七、八千年前に生きたと推定されている。
[11]
논문
Overview of genetic variation in the Y chromosome of modern Japanese males
[12]
논문
Assignment of Y-chromosomal SNPs found in Japanese population to Y-chromosomal haplogroup tree.
[13]
논문
Genetic origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages.
[14]
서적
DNA・考古・言語の学際研究が示す新・日本列島史
勉誠出版
[15]
웹사이트
https://www.23mofang[...]
[16]
문서
(この統計の対象者の出身地については通遼市232名、赤峰市212名、シリンゴル盟209名、遼寧省152名、フフホト市137名、オルドス市94名、ヒンガン盟85名、フルンボイル市82名、新疆63名、バヤンノール市49名、ウランチャブ市41名、黒竜江省38名、吉林省37名、青海省29名、包頭市25名、アルシャー盟21名、烏海市2名、その他13名となっており、著しく南東に偏っている事に留意する必要がある。)
[17]
논문
The Eurasian Heartland: A continental perspective on Y-chromosome diversity.
[18]
논문
Y-chromosomal analysis of clan structure of Kalmyks, the only European Mongol people, and their relationship to Oirat-Mongols of Inner Asia.
[19]
논문
Genomic Insights Into the Genetic Structure and Natural Selection of Mongolians.
[20]
논문
Whole-genome sequencing of 175 Mongolians uncovers population-specific genetic architecture and gene flow throughout North and East Asia.
[21]
논문
The Peopling of Korea Revealed by Analyses of Mitochondrial DNA and Y-Chromosomal Markers.
[22]
학위논문
中国西部人群的遗传混合
复旦大学
[23]
논문
Genetic substructure and admixture of Mongolians and Kazakhs inferred from genome-wide array genotyping.
[24]
논문
Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge.
[25]
논문
Genomic insights into the genetic structure and population history of Mongolians in Liaoning Province.
[26]
논문
Extensive ethnolinguistic diversity at the crossroads of North China and South Siberia reflects multiple sources of genetic diversity.
[27]
논문
Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis
[28]
논문
Genetic Relationship Between Korean and Mongolian Populations Based on the Y Chromosome DNA Variation.
[29]
논문
Genetic Diversity of Two Haploid Markers in the Udegey Population From Southeastern Siberia.
[30]
논문
Analysis of Y chromosome haplogroups in Japanese population using short amplicons and its application in forensic analysis
https://doi.org/10.1[...]
[31]
논문
High frequencies of Y-chromosome haplogroup O2b-SRY465 lineages in Korea: a genetic perspective on the peopling of Korea.
http://www.investiga[...]
[32]
논문
Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences.
2016-06
[33]
논문
スーパーサイエンスハイスクール(SSH)コンソーシアムによる日本人のY染色体とミトコンドリアDNAハプログループ地域分布調査
https://doi.org/10.1[...]
[34]
논문
Dual origins of the Japanese: Common ground for hunter-gatherer and farmer Y chromosomes
[35]
논문
Y-chromosomal binary haplogroups in the Japanese population and their relationship to 16 Y-STR polymorphisms
2007-07
[36]
문서
Mizuno 2008
[37]
논문
Lack of Association between Y-Chromosomal Haplogroups and Prostate Cancer in the Korean Population.
[38]
논문
The Origin and Composition of Korean Ethnicity Analyzed by Ancient and Present-Day Genome Sequences
[39]
간행물
Y chromosomal DNA variation in east Asian populations and its potential for inferring the peopling of Korea
https://doi.org/10.1[...]
[40]
논문
Understanding the Y chromosome variation in Korea—relevance of combined haplogroup and haplotype analyses
[41]
논문
Genomic Insights Into the Admixture History of Mongolic- and Tungusic-Speaking Populations From Southwestern East Asia
[42]
논문
The genetic structure and admixture of Manchus and Koreans in northeast China
[43]
논문
Genomic Insight Into the Population Admixture History of Tungusic-Speaking Manchu People in Northeast China
[44]
웹사이트
中国満州族集団の父系ハプログループ及び民族血統構成の状況を簡単に説明
https://www.23mofang[...]
23mofangによる
[45]
논문
Paternal Population History of East Asia: Sources, Patterns, and Microevolutionary Processes"
[46]
문서
(Table 2にある地理座標、123.2°E 40.2°Nに基づき、遼寧省[[鞍山市]][[岫巖満族自治県]]で採取されたサンプルだろうと推定できる)
[47]
논문
The Nanai Clan Samar: the Structure of Gene Pool Based on Y-Chromosome Markers
[48]
논문
Male Demography in East Asia: A North–South Contrast in Human Population Expansion Times
[49]
논문
Human paternal and maternal demographic histories: insights from high-resolution Y-chromosome and mtDNA sequences
[50]
논문
Extensive ethnolinguistic diversity at the crossroads of North China and South Siberia reflects multiple sources of genetic diversity
[51]
논문
Demographic and Genetic Portraits of the Ulchi Population
[52]
논문
Genetic Diversity of Two Haploid Markers in the Udegey Population From Southeastern Siberia
[53]
문서
"СТРУКТУРА И ФИЛОГЕОГРАФИЯ ГЕНОФОНДА КОРЕННОГО НАСЕЛЕНИЯ СИБИРИ ПО МАРКЕРАМ Y-ХРОМОСОМЫ." генетика 03.02.07
[54]
논문
The Dual Origin and Siberian Affinities of Native American Y Chromosomes
[55]
논문
Siberian genetic diversity reveals complex origins of the Samoyedic-speaking populations
[56]
논문
Investigating the Prehistory of Tungusic Peoples of Siberia and the Amur-Ussuri Region with Complete mtDNA Genome Sequences and Y-chromosomal Markers
[57]
문서
"分子人類学視野下的達斡爾族族源研究"
[58]
논문
Gene Pool of Buryats: Clinal Variability and Territorial Subdivision Based on Data of Y-Chromosome Markers
[59]
논문
Y chromosome haplotype diversity in Mongolic-speaking populations and gene conversion at the duplicated STR DYS385a,b in haplogroup C3-M407
[60]
논문
Y-chromosome diversity in the Kalmyks at the ethnical and tribal levels
[61]
논문
Genetic Diversity of the Khakass Gene Pool: Subethnic Differentiation and the Structure of Y-Chromosome Haplogroups
[62]
논문
Gene Pool Differences between Northern and Southern Altaians Inferred from the Data on Y-Chromosomal Haplogroups
[63]
논문
Estimating the impact of Mongol expansion on gene pool of Tuvans
[64]
논문
Gene-Pool Structure of Tuvinians Inferred from Y-Chromosome Marker Data
[65]
논문
Analysis of Polymorphism of Uniparental Markers in Reindeer-Herding Populations: The Tozhu Tuvans of Russia and The Tsaatans Of Mongolia
[66]
논문
Autosomal and uniparental portraits of the native populations of Sakha (Yakutia): implications for the peopling of Northeast Eurasia
http://www.biomedcen[...]
[67]
웹사이트
"СТРУКТУРА И ФИЛОГЕОГРАФИЯ ГЕНОФОНДА КОРЕННОГО НАСЕЛЕНИЯ СИБИРИ ПО МАРКЕРАМ Y-ХРОМОСОМЫ
http://www.medgeneti[...]
[68]
문서
O1b(旧O2)までしか特定されていない。論文中の系統名称は2012年時点のものであることに注意。
[69]
웹사이트
ハプログループNO (Y染色体)|NO
http://www.nature.co[...]
[70]
웹사이트
https://www.23mofang[...]
[71]
논문
A genetic landscape reshaped by recent events: Y-chromosomal insights into central Asia
https://doi.org/10.1[...]
Elsevier
[72]
논문
An updated tree of Y-chromosome Haplogroup O and revised phylogenetic positions of mutations P164 and PK4.
https://www.nature.c[...]
2011-04-20
[73]
논문
Genetic structure and demographic history of Northern Han people in Liaoning Province inferred from genome-wide array data.
[74]
논문
Developmental validation of a high-resolution panel genotyping 639 Y-chromosome SNP and InDel markers and its evolutionary features in Chinese populations.
[75]
논문
New insights into the fine-scale history of western–eastern admixture of the northwestern Chinese population in the Hexi Corridor via genome-wide genetic legacy.
[76]
논문
Fine‐scale north‐to‐south genetic admixture profile in Shaanxi Han Chinese revealed by genome‐wide demographic history reconstruction.
2022-07
[77]
논문
Forensic characteristics and genetic substructure analysis of the Handan Han population, Northern China.
[78]
논문
Multiple founding paternal lineages inferred from the newly-developed 639-plex Y-SNP panel suggested the complex admixture and migration history of Chinese people.
[79]
논문
山東漢、回族男性人群父系遺伝結構研究
2022-02
[80]
논문
Developmental validation of Y-SNP pedigree tagging system: A panel via quick ARMS PCR.
[81]
논문
The genomic history of southwestern Chinese populations demonstrated massive population migration and admixture among proto-Hmong–Mien speakers and incoming migrants
https://doi.org/10.1[...]
Springer
[82]
논문
Fine-Scale Genetic Structure and Natural Selection Signatures of Southwestern Hans Inferred From Patterns of Genome-Wide Allele, Haplotype, and Haplogroup Lineages.
[83]
논문
Taiwan Y-chromosomal DNA variation and its relationship with Island Southeast Asia
[84]
논문
Forensic Analysis and Genetic Structure Construction of Chinese Chongming Island Han Based on Y Chromosome STRs and SNPs.
[85]
논문
Genomic Insights Into the Demographic History of the Southern Chinese.
[86]
논문
Next-generation sequencing of 74 Y-SNPs to construct a concise consensus phylogeny tree for Chinese population.
2017-12
[87]
논문
Genetic Reconstruction and Forensic Analysis of Chinese Shandong and Yunnan Han Populations by Co-Analyzing Y Chromosomal STRs and SNPs.
[88]
논문
Unraveling the paternal genetic structure and forensic traits of the Hui population in Liaoning Province, China using Y-chromosome analysis.
https://doi.org/10.1[...]
[89]
웹사이트
https://www.23mofang[...]
[90]
웹사이트
23mofangによる中国イ族集団の父系ハプログループ及び民族血統構成についての簡単な紹介
https://www.23mofang[...]
[91]
논문
Retrieving Y chromosomal haplogroup trees using GWAS data.
[92]
논문
Genetic Structure of Qiangic Populations Residing in the Western Sichuan Corridor.
[93]
웹사이트
https://www.23mofang[...]
[94]
논문
The distribution of Y chromosome haplogroups in the nationalities from Yunnan Province of China.
2005-01
[95]
논문
Genetic substructure of Guizhou Tai-Kadai-speaking people inferred from genome-wide single nucleotide polymorphisms data.
[96]
논문
Fine-Scale Population Admixture Landscape of Tai–Kadai-Speaking Maonan in Southwest China Inferred From Genome-Wide SNP Data.
[97]
논문
Reconstructing the genetic admixture history of Tai-Kadai and Sinitic people: Insights from genome-wide SNP data from South China.
[98]
웹사이트
https://www.23mofang[...]
[99]
논문
The paternal genetic structure of Jingpo and Dai in southwest China.
[100]
논문
Contrasting Paternal and Maternal Genetic Histories of Thai and Lao Populations.
[101]
논문
Cultural variation impacts paternal and maternal genetic lineages of the Hmong-Mien and Sino-Tibetan groups from Thailand.
[102]
논문
Y chromosomal evidence on the origin of northern Thai people.
[103]
논문
The paternal and maternal genetic history of Vietnamese populations.
[104]
논문
Patrilineal Perspective on the Austronesian Diffusion in Mainland Southeast Asia.
[105]
논문
Major East–West Division Underlies Y Chromosome Stratification across Indonesia.
[106]
논문
"The Y-chromosome landscape of the Philippines: extensive heterogeneity and varying genetic affinities of Negrito and non-Negrito groups."
2011
[107]
논문
"The Dual Origin of the Malagasy in Island Southeast Asia and East Africa: Evidence from Maternal and Paternal Lineages."
2005
[108]
논문
"Isolation, contact and social behavior shaped genetic diversity in West Timor."
2014
[109]
논문
"Reduced Y-Chromosome, but Not Mitochondrial DNA, Diversity in Human Populations from West New Guinea."
2003
[110]
논문
"Unexpected NRY Chromosome Variation in Northern Island Melanesia."
2006
[111]
논문
"Melanesian and Asian Origins of Polynesians: mtDNA and Y Chromosome Gradients Across the Pacific."
2006
[112]
논문
"Genetic Evidence for Modifying Oceanic Boundaries Relative to Fiji."
2016
[113]
논문
"Bridging Near and Remote Oceania: mtDNA and NRY Variation in the Solomon Islands."
2012
[114]
웹사이트
https://www.23mofang[...]
[115]
웹사이트
ハプログループO1b2-P49の系統樹
https://www.theytree[...]
TheYtree
[116]
웹사이트
https://discover.fam[...]
[117]
웹사이트
https://discover.fam[...]
[118]
논문
"Y-SNP miniplexes for East Asian Y-chromosomal haplogroup determination in degraded DNA."
2013
[119]
웹사이트
https://www.23mofang[...]
[120]
웹사이트
ハプログループO-P49の系統樹
https://www.yfull.co[...]
YFull
[121]
논문
"Genetic Relationship Between Korean and Mongolian Populations Based on the Y Chromosome DNA Variation."
2003
[122]
웹사이트
https://korean.theyt[...]
[123]
논문
"Multiple Human Population Movements and Cultural Dispersal Events Shaped the Landscape of Chinese Paternal Heritage"
2024-07-01
[124]
웹사이트
https://discover.fam[...]
[125]
웹사이트
https://discover.fam[...]
[126]
웹사이트
https://www.23mofang[...]
[127]
논문
"An update and frequency distribution of Y chromosome haplogroups in modern Japanese males."
2023
[128]
웹사이트
https://www.23mofang[...]
[129]
웹사이트
2019-2020 Haplogroup O Tree
https://docs.google.[...]
[130]
웹사이트
Overview of genetic variation in the Y chromosome of modern Japanese males
https://www.jstage.j[...]
[131]
웹사이트
An update and frequency distribution of Y chromosome haplogroups in modern Japanese males
https://www.nature.c[...]
[132]
웹사이트
https://discover.fam[...]
[133]
웹사이트
https://discover.fam[...]
[134]
웹사이트
https://discover.fam[...]
[135]
웹사이트
https://www.23mofang[...]
[136]
웹사이트
https://www.23mofang[...]
[137]
웹사이트
https://discover.fam[...]
[138]
웹사이트
https://www.23mofang[...]
[139]
웹사이트
https://www.23mofang[...]
[140]
웹사이트
https://discover.fam[...]
[141]
웹사이트
https://www.23mofang[...]
[142]
웹사이트
https://discover.fam[...]
[143]
웹사이트
https://www.23mofang[...]
[144]
웹사이트
FamilyTreeDNA Discover™ Time Tree for Haplogroup O-P49
https://discover.fam[...]
[145]
웹사이트
https://www.23mofang[...]
[146]
논문
Y Chromosomes of 40% Chinese Descend from Three Neolithic Super-Grandfathers.
[147]
웹사이트
23魔方祖源による「豫陕晋冀王氏家族」の説明
https://www.23mofang[...]
[148]
웹사이트
Phylogenetic tree of Haplogroup O-P49 at 23mofang
https://www.23mofang[...]
[149]
웹사이트
23魔方祖源による「湖南周氏家族」の説明
https://www.23mofang[...]
[150]
웹사이트
http://forensic.yons[...]
[151]
웹사이트
https://discover.fam[...]
[152]
논문
Confirmation of Y haplogroup tree topologies with newly suggested Y-SNPs for the C2, O2b and O3a subhaplogroups.
[153]
웹사이트
https://www.23mofang[...]
[154]
웹사이트
https://discover.fam[...]
[155]
웹사이트
https://www.theytree[...]
[156]
웹사이트
ロシア極東新石器時代研究の新展開
http://21coe.kokugak[...]
[157]
간행물
[調査研究活動報告] 鳥取県鳥取市青谷上寺地遺跡出土弥生後期人骨のDNA分析
https://rekihaku.rep[...]
国立歴史民俗博物館
2020-03
[158]
웹사이트
Phylogenetic tree of O-CTS713 at TheYtree
https://www.theytree[...]
2023-04-27
[159]
간행물
西北九州弥生人の遺伝的な特徴
https://doi.org/10.1[...]
日本人類学会
[160]
간행물
[調査研究活動報告]鳥取県鳥取市青谷上寺地遺跡出土弥生後期人骨の核DNA分析
https://rekihaku.rep[...]
国立歴史民俗博物館
2024-01-18
[161]
웹사이트
YFull
http://www.yfull.com[...]
[162]
문서
(Xue 2006), (Katoh 2004), (Jin 2009), (Nonaka 2007), (Poznik 2016), and (Hammer 2006)
[163]
문서
(Hammer 2006), (Xue 2006), (Katoh 2004), (Kim 2007), and (Park 2013)
[164]
블로그
https://blog.naver.c[...]
[165]
문서
(Hammer 2006) and (Nonaka 2007)
[166]
문서
(Karafet 2001), (Jin 2003), (Katoh 2004), and (Xue 2006)
[167]
블로그
https://blog.naver.c[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com